Research
Our laboratory has diverse research interests that span microbial signaling, evolution, antimicrobial development, to the development of immunomodulatory therapeutics.
1. Elucidating the c-di-GMP signal transduction pathway in V. cholerae. A major focus of our laboratory is to use V. cholerae as a model system to discover fundamental principles about c-di-GMP signaling. To this end, we have shown that two transcription factors central in the regulation of biofilm formation and motility directly bind c-di-GMP. VpsR, the transcriptional activator of biofilm formation, is activated upon c-di-GMP binding while FlrA, the master regulator of motility, is inhibited. This results in an opposing regulatory pathway in which c-di-GMP acts as both a co- and anti-activator. Building on these studies we developed the first in vitro transcription system to show that c-di-GMP binding to VpsR drives it to induce open complex formation in RNA polymerase. A systematic analysis of c-di-GMP signaling in V. cholerae revealed high-specificity signaling indicating that c-di-GMP produced from different synthesis enzymes differentially impacts downstream phenotypes. We have also identified environmental factors that control c-di-GMP signaling and discovered that the intestinal cues bile and bicarbonate inversely control c-di-GMP levels, likely providing V. cholerae spatial cues within the human host. We have also demonstrated that the c-di-GMP-binding riboswitch Vc2 controls motility via inhibition of the downstream transcription factor TfoY. Finally, we have demonstrated a novel mechanism of gene regulation showing that Vc2 stabilizes a novel sRNA in cis by preventing its 3’ degradation. Below is a simplified model of the inverse regulation of biofilm formation and motility by c-di-GMP in V. cholerae.

2. Expanding our understanding of phenotypes regulated by c-di-GMP in V. cholerae. It has long been appreciated that c-di-GMP positively regulates biofilms and negatively impacts motility. However, it is known that the regulation of hundreds of genes changes when c-di-GMP levels are increased. We have recently discovered that c-di-GMP regulates numerous other phenotypes in V. cholerae including Type II Secretion, DNA repair, and stress responses to hydrogen peroxide. Furthermore, all of these traits are regulated as part of the VpsR/VpsT regulatory network, which also induces the genes necessary for biofilm formation, suggesting that these additional phenotypes lead to increased fitness of V. cholerae within a biofilm. Finally, we have shown that c-di-GMP also regulates cell curvature of V. cholerae. This cartoon shows a partial model of our understanding of c-di-GMP regulated phenotypes in V. cholerae.

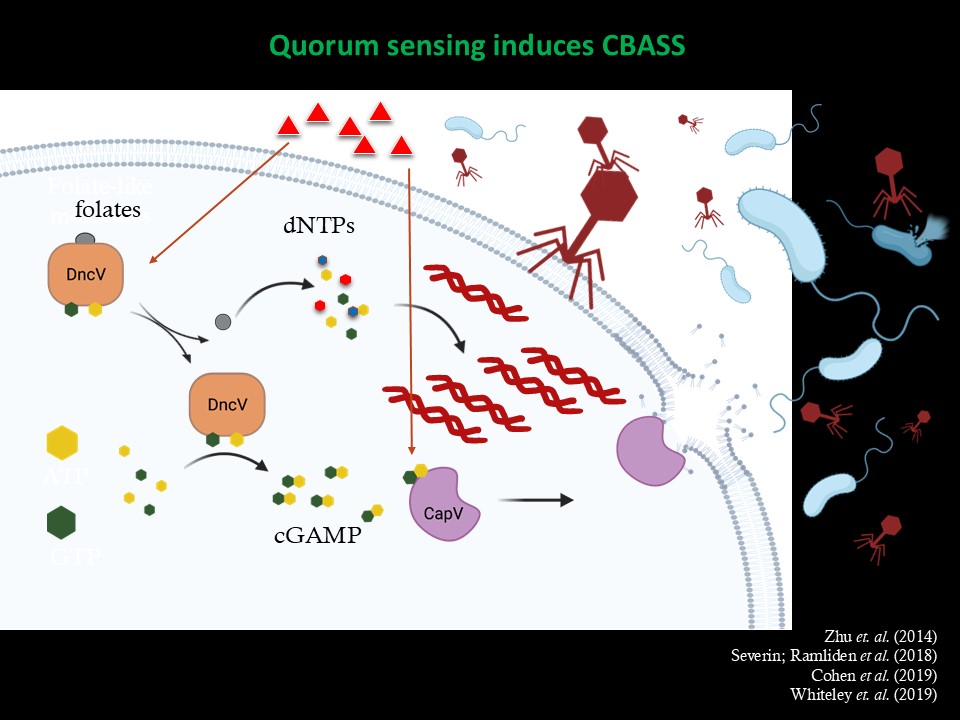
3. Discovery of the first cyclic GMP-AMP signaling pathway in bacteria. The newest cyclic di-nucleotide to be discovered in bacteria is the signaling molecule cyclic GMP-AMP in 2012. This signal was shown to be synthesized by the enzyme DncV encoded on a genomic island that is specific to the 7th, and current, pandemic of V. cholerae. We recently found that cGAMP synthesized by DncV activates a putative unknown phospholipase, which we named CapV, to remodel the membrane of V. cholerae. CapV is found immediately 5’ of DncV on this genomic island, suggesting these genes are a functional cassette that can move by horizontal gene transfer. This work has identified the first ever bacterial protein receptor of cGAMP and has significant implications on the evolution of V. cholerae pandemics. In the cartoon below, El Tor V. cholerae is represented by the hot dog, and its acquisition of two unique genomic islands, VSP-1 and VSP-2, by ketchup and mustard. dncV and capV are both encoded in VSP-1. cGAMP and CapV are the founding members of a widespread phage defense system known as CBASS.
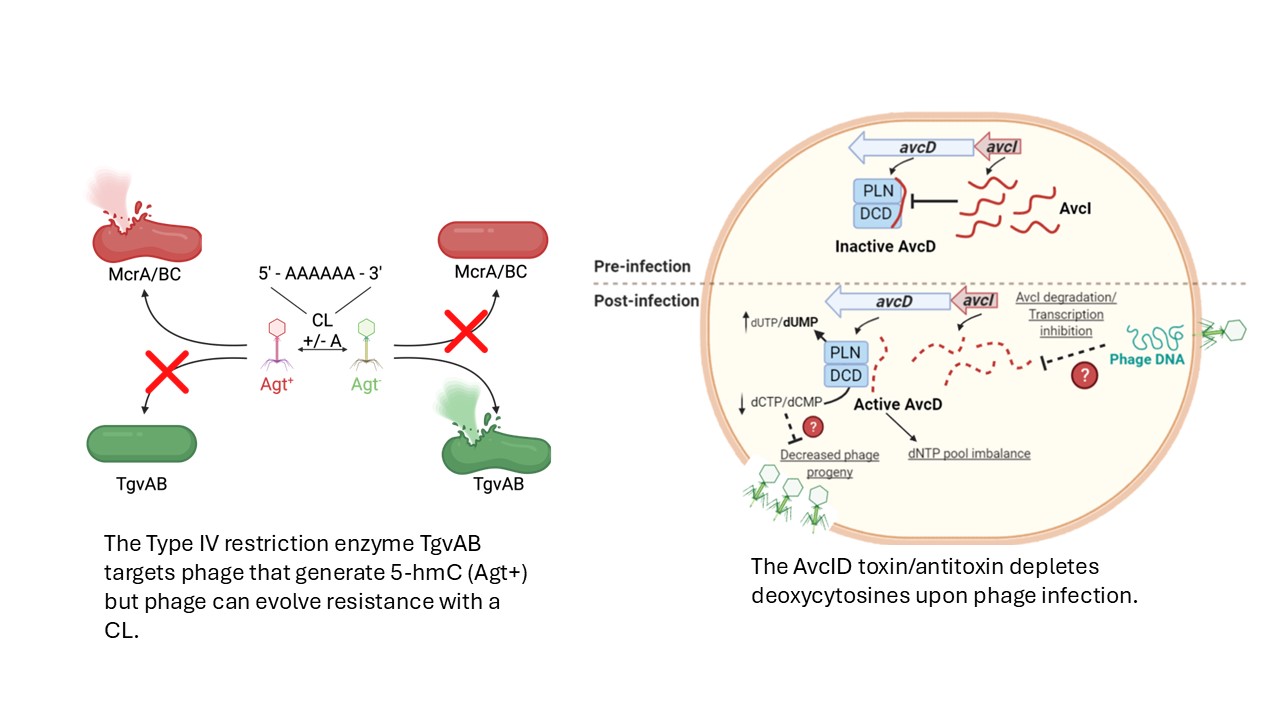
4. Discovering new phage defense and understanding phage evolution. Inspired by our discovery of CapV, we have used Vibrio cholerae as a model system to discover several other phage defense systems including AvcID, a toxin-antitoxin that depletes cellular deoxycytosines, and TgvAB, a Type IV restirction enzyme that restrict phages that possess 5-hydroxymethylcytosine modified DNA genomes. These discoveries led us to discovery that phages can overcome these defense using highly mutagenic single base repeats in the genome using a bet-hedging strategy. These repeats are known as contigency loci, and our work is the first to describe these in phages!